|
|
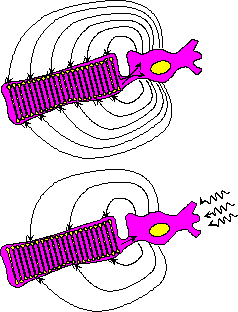
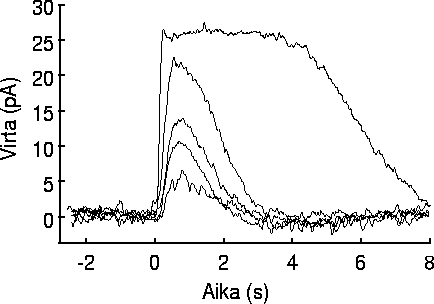
| (a) Valon säätelemä pimeävirta on sähköisesti mitattavissa oleva suure, joka riippuu reseptorin absorboimasta valomäärästä. | (b) Salamanterin sauvasolusta mitattu virtasignaali, kun soluun on kohdistettu valosalamia eri intensiteeteillä. |
Sauvasolussa kiertää pimeässä Na+-ioneiden kuljettama
sähkövirta.![[*]](foot_motif.gif) Valon vaikutuksesta tämä ns. pimeävirta
pienenee (kuva 2.3(a)). Normaalin hermosolun tapaan
sauvasolu pyrkii aktiivisesti pitämään Na+-ionien konsentraation
solun sisällä alhaisena. Tästä huolehtivat Na+/K+
-pumppumolekyylit, jotka sijaitsevat (pääasiassa) solun sisäjäsenessä.
ATP-energian avulla nämä entsyymit siirtävät Na+-ioneja solun
ulkopuolelle ja K+-ioneja solun sisään. Näin Na+-ioneille muodostuu
sähkökemiallinen gradientti, joka ajaa niitä solun sisään.
Na+-ionit pääsevät virtaamaan takaisin soluun ulkojäsenen
kationikanavien kautta. Solussa kulkee siis
Na+-ionien kuljettama sähkövirta sisäjäsenestä solun ulkoiseen
tilaan, sieltä ulkojäsenen kationikanavien kautta ulkojäseneen ja
limasillan kautta takaisin sisäjäseneen. Valon vaikutuksesta
ulkojäsenen kationikanavia sulkeutuu ja tämä virta pienenee.
Valon vaikutuksesta tämä ns. pimeävirta
pienenee (kuva 2.3(a)). Normaalin hermosolun tapaan
sauvasolu pyrkii aktiivisesti pitämään Na+-ionien konsentraation
solun sisällä alhaisena. Tästä huolehtivat Na+/K+
-pumppumolekyylit, jotka sijaitsevat (pääasiassa) solun sisäjäsenessä.
ATP-energian avulla nämä entsyymit siirtävät Na+-ioneja solun
ulkopuolelle ja K+-ioneja solun sisään. Näin Na+-ioneille muodostuu
sähkökemiallinen gradientti, joka ajaa niitä solun sisään.
Na+-ionit pääsevät virtaamaan takaisin soluun ulkojäsenen
kationikanavien kautta. Solussa kulkee siis
Na+-ionien kuljettama sähkövirta sisäjäsenestä solun ulkoiseen
tilaan, sieltä ulkojäsenen kationikanavien kautta ulkojäseneen ja
limasillan kautta takaisin sisäjäseneen. Valon vaikutuksesta
ulkojäsenen kationikanavia sulkeutuu ja tämä virta pienenee.
Näköreseptorin ulkojäsenen kalvokiekkojen näköpigmentti muodostaa
ensimmäisen linkin valoinformaation muuttumisessa biosähköiseksi
signaaliksi. Näköpigmentti määrää solun havaitseman valon
aallonpituuden. Tappijärjestelmän värien erottelu perustuu eri
tappityyppien pigmenttien erilaiseen absorptiospektriin. Esimerkiksi
ihmisen kolmen tappityypin absorptiomaksimit ovat ![]() ,
, ![]() ja
ja ![]() [3].
Yleisesti puhutaan sini-, viher- ja punaherkistä tapeista niiden
absorboiman valon värin mukaan. Sauvasolun
pigmenttiproteiinia, rodopsiinia, kutsutaan näköpurppuraksi pigmentin
punertavan värin mukaan. Toisin kuin tappien tapauksessa, puhutaan
usein punaisista sauvoista, vaikka sauvojen absorptiomaksimi
[3].
Yleisesti puhutaan sini-, viher- ja punaherkistä tapeista niiden
absorboiman valon värin mukaan. Sauvasolun
pigmenttiproteiinia, rodopsiinia, kutsutaan näköpurppuraksi pigmentin
punertavan värin mukaan. Toisin kuin tappien tapauksessa, puhutaan
usein punaisista sauvoista, vaikka sauvojen absorptiomaksimi
![]() onkin sinivihreän valon aallonpituuden kohdalla
[4].
onkin sinivihreän valon aallonpituuden kohdalla
[4].
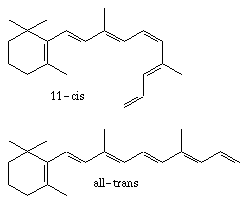
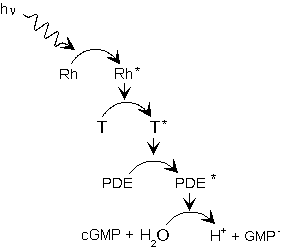
Rodopsiinin fotoneita absorboiva osa on rodopsiinin proteiiniosaan, opsiiniin, kovalentisti sitoutunut retinaali-molekyyli, joka on A-vitamiinin aldehydi (kuva 2.4(a)). Fotonin absorptio aiheuttaa retinaalin fotoisomerisaation, joka johtaa myös opsiinin konformiseen muodonmuutokseen. Kuvassa 2.4(b) on esitetty fotonin absorptiosta ja rodopsiinin konformaatiomuutoksesta alkava kemiallinen ketjureaktio, joka johtaa lopulta solunsisäisen syklisen guanosiinimonofosfaatin (cGMP) konsentraation laskuun ja solukalvon cGMP-herkkien kationikanavien sulkeutumiseen. Fotonin absorboinut aktivoitunut rodopsiini Rh* aktivoi suuren joukon G-proteiineja (sauvassa transdusiini), joista kukin aktivoi yhden PDE-entsyymimolekyylin. PDE hajottaa puolestaan cGMP:tä, joka pitää sauvasolun ulkojäsenen solukalvon cGMP-herkkiä kationikanavia avoimina. Kanava päästää lävitseen Na+-ioneja, jos kanavaan on sitoutunut kolme cGMP-molekyyliä. Kun cGMP:n konsentraatio laskee, kanavia sulkeutuu ja Na+-ionien pääsy soluun estyy.
|
|
| (a) Retinaalin fotoisomerisaatiosa kaksoissidoksia sisältävä hiiliketju oikenee. | (b) Fotonin laukaisema ketjureaktio johtaa cGMP:n konsentraation laskuun. Kaavion symbolit: Rh = rodopsiini, T =transdusiini, PDE =fosfodiesteraasi, cGMP =syklinen guanosiinimonofosfaatti. |
Sauvasolun pimeävirta on sähköisesti mitattavissa oleva suure, minkä vuoksi sauvasolu on varsin hyvä tutkimuskohde. Koska virta on valon säätelemä, sauvasolun tilaa voidaan tutkia tarkkailemalla sauvaan kohdistuvan valon intensiteetin ja pimeävirran muodostaman stimulus/vaste -parin suhdetta. Yleensä mitataan virran muutosta, kun sauvaan kohdistetaan lyhyt valosalama, jonka intensiteettiä voidaan vaihdella. Kuvassa 2.3(b) on esitetty salamanterin sauvasolusta mitattu virta, kun sauvaan on kohdistettu valosalamia eri intensiteeteillä. Kuvaajassa ajan nollakohtana on käytetty hetkeä, jolloin sauvaan on kohdistettu 20 ms valosalama. Pystyakselin nollataso vastaa tilannetta pimeässä (kanavia auki) ja pystyakselin positiiviset arvot kuvaavat pimeävirran laskua (kanavat sulkeutuvat). Tätä merkkikäytäntöä käytetään usein, koska tarkastelun kohteena oleva suure on valon aiheuttama muutos pimeävirrassa eikä pimeävirran taso sinänsä. Valovastesignaalin luonne näkyy hyvin kuvaajista. Pienellä intensiteetillä vaste nousee maksimiinsa hitaammin kuin suurella intensiteetillä. Myös vasteen maksimiamplitudi kasvaa intensiteetin kasvaessa kunnes intensiteetti on kasvanut tasolle, jolla valosalama aiheuttaa solun kaikkien Na+-kanavien sulkeutumisen. Jos valosalaman intensiteettiä tästä vielä kasvatetaan, vasteen maksimiamplitudi pysyy samana, mutta vasteen kesto pitenee eli Na+-kanavat pysyvät pidempään suljettuina.