Kuvassa 2.2 on esitetty kaaviomaisesti selkärankaisen
silmän, verkkokalvon ja näköreseptorisolun rakenne. Kuvan mukainen
rakenne pätee esimerkiksi ihmisen silmään. Silmän optinen rakenne on
kameran kaltainen. Linssi, jonka polttoväliä eliö pystyy säätelemään,
projisoi ympäristön kuvan verkkokalvolle, joka vastaa kameran
valotettavaa filmiä. Verkkokalvo on kiinnittynyt silmän pohjan
pigmenttiepiteeliin, joka päiväeläimillä on tyypillisesti musta. Musta
pigmenttiepiteeli vähentää heijastuksia ja parantaa siten erotuskykyä.
Toisaalta monet yöeläimet pyrkivät parantamaan näön herkkyyttä
heijastavalla pigmenttiepiteelillä.![[*]](foot_motif.gif)
Itse verkkokalvon rakenne on pääpiirteissään kolmikerroksinen. Pigmenttiepiteeliin kiinnittyneinä ovat näköreseptorit, jotka absorboivat valokvantteja ja muuttavat valoinformaation sähköiseksi signaaliksi. Reseptorisolut kytkeytyvät synapseilla bipolaarisoluihin, jotka muodostavat verkkokalvon toisen kerroksen. Reseptorisolujen ja bipolaarisolujen väliset kytkennät voivat olla joko ärsyttäviä tai ehkäiseviä. Sekä reseptorisolujen, että bipolaarisolujen sähköinen signaali on luonteeltaan hidas reseptoripotentiaali. Bipolaarisolut kytkeytyvät puolestaan gangliosoluihin. Tällä tasolla synapsit ovat vain ärsyttäviä. Gangliosolusta lähtevä signaali on pulssimainen aktiopotentiaali, joka kulkee solun aksonia pitkin verkkokalvon päällä silmän sokeaan täplään, josta aksonit jatkavat näköhermona ja välittävät informaation edelleen korkeamman tason hermosoluille. Verkkokalvossa on myös poikittais- ja takaisinkytkentöjä muodostavia amakriini- ja horisontaalisoluja sekä ns. glia-solukkoa.
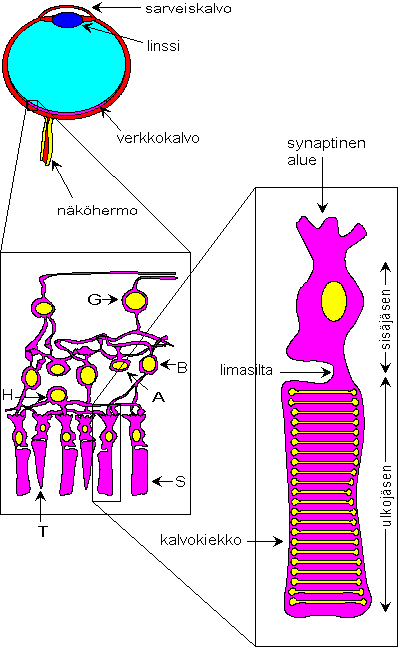 |
Reseptorisolujen rakenne on kolmiosainen. Lähimpänä pigmenttiepiteeliä on solun ulkojäsen. Ulkojäsen kytkeytyy ohuella limasillalla sisäjäseneen. Kolmannen osan muodostaa solun synaptinen alue, joka voi myös olla varsin ohuella haarakkeella kiinni sisäjäsenessä. Ulkojäsen koostuu pääasiassa ns. kiekkokalvostosta, joka sisältää valokvantteja absorboivaa pigmenttiä. Sisäjäsenessä sijaitsevat mm. solun tuma ja mitokondriot. Synaptinen alue on nimensä mukaisesti se solun osa, joka muodostaa synapseja muihin hermosoluihin.
Selkärankaisilla reseptorisolut voidaan jakaa kahteen päätyyppiin. Solutyypit eroavat toisistaan sekä ulkonäkönsä että toiminnallisten ominaisuuksien suhteen. Esimerkiksi sammakolla tappisolun ulkojäsen on kartiomainen ja lyhyt kun taas sauvasolun ulkojäsen on sylinterimäinen ja pitkä. Myös solutyyppien kiekkokalvoston rakenne on erilainen. Sauvasolussa kalvosto muodostuu irrallisista kiekoista ulkojäsenen solukalvon sisällä. Tappisolussa kiekkokalvosto muodostuu laskostuneesta ulkojäsenen solukalvosta. Reseptorityyppien ulkonäölliset erot eivät ole kaikilla selkärankaisilla yhtä selkeitä. Esimerkiksi ihmisen tapit ja sauvat ovat lähes yhtä pitkiä.
Toiminnallisesti tapit vastaavat värinäöstä ja tarkkojen yksityiskohtien havainnoinnista näkökentän keskellä hyvässä valaistuksessa. Tapit myös toipuvat nopeasti valostimulaatiosta, joten tappijärjestelmällä on hyvä temporaalinen erotuskyky. Sauvat ovat puolestaan erikoistuneet hämäränäköön ja näkökentän reunaosiin. Tämä työjako näkyy solujen sijoittumisessa verkkokalvolle. Esimerkiksi ihmisen verkkokalvon keskiosassa ns. keltaisessa täplässä on vain tappisoluja. Tämä tarkan näön alue on halkaisijaltaan noin 0.2 mm ja näkökentästä se kattaa noin yhden asteen kulman [3]. Siirryttäessä verkokalvon keskeltä kohti periferiaa tappitiheys pienenee ja sauvojen osuus reseptoreista kasvaa. Myös reseptoreiden kytkeytyminen korkeamman tason hermosoluihin muuttuu. Tarkan näön alueella kukin tappi on kytkeytynyt vain yhteen gangliosoluun. Siirryttäessä verkkokalvon reuna-alueita kohti gangliosolujen reseptiivinen kenttä kasvaa, eli ne keräävät informaatiota useammalta reseptorilta ja laajemmalta alueelta. Tämän seurauksena näön tarkkuus verkkokalvon reunaosissa on huono, mutta samalla saavutetaan erittäin herkkä pientenkin valaistusmuutosten havainnointikyky, minkä seurauksena esimerkiksi liikkeet näkökentän laidalla havaitaan tehokkaasti.